細胞因子信號轉導
正如在“細胞因子的功能是什么?”的文章中提到的,細胞因子是一種調節多種生物功能的分泌蛋白。它們通過表達在細胞表面的特異性受體來激活復雜的信號轉導通路,包括Janus激酶信號轉導和轉錄激活劑(JAK-STAT)通路、胰島素受體底物(IRS)蛋白、磷脂酰肌醇3激酶(PI3K)、Ras/mitogen激活蛋白(MAP)激酶級聯等,發揮其生物功能。其中,JAK-STAT通路是目前研究較多的,也是主流。本文主要針對JAK-STAT通路及相關細胞因子進行研究。
參與JAK-STAT途徑的細胞因子
細胞因子對大多數細胞過程的調控,往往是一致的,而且是通過類似的信號轉導途徑。JAK-STAT信號轉導途徑是I型細胞因子和干擾素參與的主要信號轉導途徑,干擾素也被稱為II型細胞因子。總體而言,細胞因子大致可分為I型和II型細胞因子。
I型與II型細胞因子的命名是基于細胞因子及其受體的三維結構。I型細胞因子一般具有“上-上-下”的4α-螺旋結構。I型細胞因子代表了一個龐大的蛋白質家族,包括許多由一種白細胞產生并作用于另一種白細胞的白細胞介素(ILs),比如IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-12、IL-13、IL-15和IL-21。除這些ILs外,I型細胞因子還包括胸腺基質淋巴細胞生成素(TSLP)、SCF、GCSF、GMCSF、紅細胞生成素(EPO)、血小板生成素(TPO)、LIF、抑瘤素M(OSM)、人心肌營養素1(CT-1)、睫狀神經營養因子(CNTF);以及其他重要的蛋白質,包括瘦素(Leptin)、生長激素和催乳素(Prolactin)。
II型細胞因子指I型干擾素(IFN-α/β)和II型干擾素(IFN-γ)以及IL-10和IL-22。盡管它們被叫做為白細胞介素,但比起與I型細胞因子的聯系,它們與干擾素的關系更為緊密。
細胞因子激活的JAK-STAT信號通路機制
如圖1所示,當細胞因子與其細胞表面的受體結合時,促進了JAK-STAT信號激活,導致受體的二聚化和受體相關的JAKs的激活。激活的JAK蛋白使受體磷酸化,為STATs創造結合位點,STATs也被JAK磷酸化。一旦被磷酸化,STAT蛋白二聚化,在運輸到細胞核后誘導細胞因子所調節的下游基因的轉錄。相反,信號轉導能被以下三種不同的機制所抑制。SHP-1、PIAS和SOCS都是組成性表達基因。SHP-1可使活化的JAKs或受體去磷酸化。PIAS可使STATs蛋白SUMO化修飾,抑制STATs轉錄激活。SOCS在細胞因子信號傳導的作用下被激活,可以抑制JAK的活性或靶向信號傳導分子的泛素化和蛋白溶解。
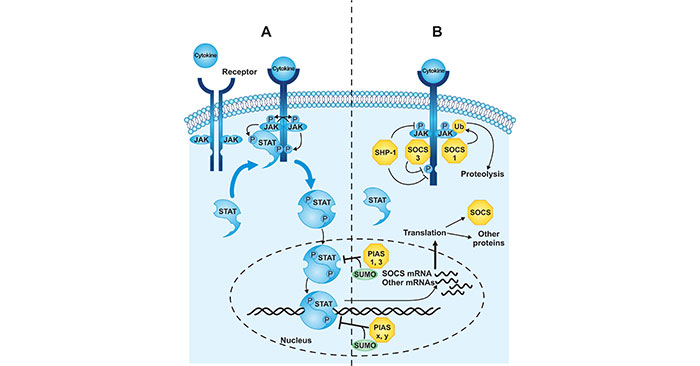
圖1. JAK-STAT信號轉導途徑示意圖
*圖片來源于J Biol Chem的出版物 [1]
在JAK-STAT信號通路中,細胞因子激活的JAKs和STATs不同。例如,大多數共享γc的I型細胞因子激活的JAKs為Jak1、Jak3,如IL-2、IL-7、IL-9、IL-15和IL-4;激活的STATs為Stat5a、Stat5b、Stat3和Stat6。而共享βc的I型細胞因子(IL-3、IL-5、GM-CSF)激活的JAKs是Jak2;激活的STATs為Stat5a和Stat5b。此外,以II型細胞因子IFN-α/β為例,激活的JAKs包括Jak1和Tyk2;激活的STATs包括Stat1和Stat2。
參考文獻:
[1] Wormald Samuel,Hilton Douglas J,Inhibitors of cytokine signal transduction.[J] .J Biol Chem, 2004, 279: 821-4.





