-
中文名稱:人絲氨酸/精氨酸富集剪接因子1(SFRS1)酶聯免疫試劑盒
-
貨號:CSB-EL021142HU
-
規格:96T/48T
-
價格:¥3600/¥2500
-
其他:

產品詳情
-
產品描述:人絲氨酸/精氨酸富集剪接因子1(SFRS1)酶聯免疫試劑盒(CSB-EL021142HU)為雙抗夾心法ELISA試劑盒,定量檢測血清、血漿、組織勻漿、細胞裂解物樣本中的SFRS1含量。SFRS1(絲氨酸/精氨酸富集剪接因子1)是一種調控RNA剪接的關鍵因子,與多種癌癥的發生發展密切相關。其通過調控VEGFA mRNA的可變剪接,促進食管鱗狀細胞癌細胞的增殖、侵襲和遷移。試劑盒檢測范圍為31.25 pg/mL-2000 pg/mL,適用于基礎科研中SFRS1功能研究,如腫瘤微環境調控、神經退行性疾病模型或細胞應激反應中的剪接機制探索,為分子生物學、病理學及藥物開發等領域的蛋白表達分析提供可靠工具。本品僅用于科研,不用于臨床診斷,產品具體參數及操作步驟詳見產品說明書。
-
別名:Alternative splicing factor 1 ELISA Kit; Alternative-splicing factor 1 ELISA Kit; arginine/serine-rich 1 ELISA Kit; ASF 1 ELISA Kit; ASF ELISA Kit; ASF-1 ELISA Kit; ASF1 ELISA Kit; FLJ53078 ELISA Kit; MGC5228 ELISA Kit; P33 subunit ELISA Kit; Pre mRNA splicing factor SF2 P33 subunit ELISA Kit; pre-mRNA-splicing factor SF2 ELISA Kit; Serine/arginine-rich splicing factor 1 ELISA Kit; SF2 ELISA Kit; SF2P33 ELISA Kit; SFRS1 ELISA Kit; Splicing factor 2 alternate splicing factor ELISA Kit; Splicing factor 2 ELISA Kit; Splicing factor ELISA Kit; Splicing factor arginine/serine rich 1 ELISA Kit; SR Splicing factor 1 ELISA Kit; SRp30a ELISA Kit; srsf1 ELISA Kit; SRSF1_HUMAN ELISA Kit
-
縮寫:SFRS1
-
Uniprot No.:
-
種屬:Homo sapiens (Human)
-
樣本類型:serum, plasma, tissue homogenates, cell lysates
-
檢測范圍:31.25 pg/mL-2000 pg/mL
-
靈敏度:7.81 pg/mL
-
反應時間:1-5h
-
樣本體積:50-100ul
-
檢測波長:450 nm
-
研究領域:Epigenetics and Nuclear Signaling
-
測定原理:quantitative
-
測定方法:Sandwich
-
精密度:
Intra-assay Precision (Precision within an assay): CV%<8% Three samples of known concentration were tested twenty times on one plate to assess. Inter-assay Precision (Precision between assays): CV%<10% Three samples of known concentration were tested in twenty assays to assess. -
線性度:
To assess the linearity of the assay, samples were spiked with high concentrations of human SFRS1 in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. Sample Serum(n=4) 1:1 Average % 93 Range % 88-97 1:2 Average % 96 Range % 90-101 1:4 Average % 99 Range % 96-104 1:8 Average % 102 Range % 96-106 -
回收率:
The recovery of human SFRS1 spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. Sample Type Average % Recovery Range Serum (n=5) 92 89-98 EDTA plasma (n=4) 98 95-103 -
標準曲線:
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. 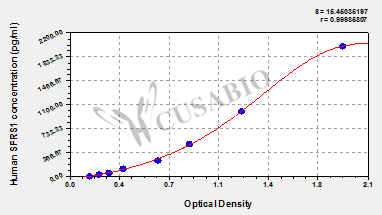
pg/ml OD1 OD2 Average Corrected 2000 1.875 1.969 1.922 1.766 1000 1.196 1.232 1.214 1.058 500 0.823 0.877 0.850 0.694 250 0.635 0.625 0.630 0.474 125 0.394 0.385 0.390 0.234 62.5 0.287 0.291 0.289 0.133 31.25 0.215 0.222 0.219 0.063 0 0.161 0.151 0.156 -
數據處理:
-
貨期:3-5 working days
引用文獻
- Single Cell Immuno-laser Microdissection Coupled to Label-free Proteomics to Reveal the Proteotypes of Human Brain Cells After Ischemia Garca-Berrocoso T.et al,Mol Cell Proteomics,2017
產品評價

相關產品
靶點詳情
-
功能:Plays a role in preventing exon skipping, ensuring the accuracy of splicing and regulating alternative splicing. Interacts with other spliceosomal components, via the RS domains, to form a bridge between the 5'- and 3'-splice site binding components, U1 snRNP and U2AF. Can stimulate binding of U1 snRNP to a 5'-splice site-containing pre-mRNA. Binds to purine-rich RNA sequences, either the octamer, 5'-RGAAGAAC-3' (r=A or G) or the decamers, AGGACAGAGC/AGGACGAAGC. Binds preferentially to the 5'-CGAGGCG-3' motif in vitro. Three copies of the octamer constitute a powerful splicing enhancer in vitro, the ASF/SF2 splicing enhancer (ASE) which can specifically activate ASE-dependent splicing. Isoform ASF-2 and isoform ASF-3 act as splicing repressors. May function as export adapter involved in mRNA nuclear export through the TAP/NXF1 pathway.
-
基因功能參考文獻:
- SRSF1 promotes vascular smooth muscle cell (VSMC) proliferation and injury-induced neointima formation. SRSF1 favors the induction of a truncated p53 isoform, Delta133p53, which has an equal proliferative effect and in turn transcriptionally activates Kruppel-like factor 5 (KLF5) via the Delta133p53-EGR1 complex, resulting in an accelerated cell-cycle progression and increased VSMC proliferation. PMID: 28799539
- repeat RNA-sequestration of SRSF1 triggers the NXF1-dependent nuclear export of C9ORF72 transcripts retaining expanded hexanucleotide repeats PMID: 28677678
- A -44 G to A "hot zone" putative functional noncoding variant of SRSF1 was found in an AML patient. It alters the binding activities of E2F6, ELF1, and ELK4, ELK4. PMID: 29764005
- We now show that the ability of SRPK1 to mobilize SRSF1 from speckles to the nucleoplasm is dependent on active CLK1. Diffusion from speckles is promoted by the formation of an SRPK1-CLK1 complex that facilitates dissociation of SRSF1 from CLK1 and enhances the phosphorylation of several serine-proline dipeptides in this SR protein PMID: 29335301
- Authors showed that Mir505-3p was capable of inhibiting tumor proliferation driven by SRSF1 in two neural tumor cell lines, Neuro-2a (N2a) and U251, exclusively in serum-reduced condition. Authors observed that the protein level of SRSF1 was gradually promoted by increasing concentration of serum. PMID: 29120871
- The present study suggested that the tumor suppressor miR30c may be involved in prostate cancer tumorigenesis, possibly via targeting ASF/SF2. PMID: 28677791
- It has been proposed that SF2/ASF has a protective role against JC virus reactivation in multiple sclerosis patients. PMID: 27812788
- Immune suppression of JC virus gene expression is mediated by SRSF1. PMID: 26951564
- ASF/SF2 is identified as a splicing regulator of cyclin T1, which contributes to the control of the subsequent transcription events. PMID: 28422315
- Findings suggest MALAT1 increases AKAP-9 expression by promoting SRPK1-catalyzed SRSF1 phosphorylation in CRC cells. These results reveal a novel molecular mechanism by which MALAT1 regulates AKAP-9 expression in CRC cells. PMID: 26887056
- high level of SF2, as a novel oncoprotein in RCC, was significantly associated with poor survival in a large cohort of RCC specimens. Taken together, our study presents a road map for the prediction and validation of miR-766-3p/SF2 axis and thus imparts a therapeutic way for further RCC progression. PMID: 28657135
- We present a joint atomistic molecular dynamics (MD) and experimental study of two RRM-containing proteins bound with their single-stranded target RNAs, namely the Fox-1 and SRSF1 complexes.The simulations predict unanticipated specific participation of Arg142 at the protein-RNA interface of the SRFS1 complex, which is subsequently confirmed by NMR and ITC measurements PMID: 27193998
- Using NMR spectroscopy with two separately expressed domains of SRSF1, we showed that several residues in the RNA-binding motif 2 interact with the N-terminal region of the RS domain (RS1). PMID: 27091468
- Especially, in SRSF1 morphants, bone cartilage formation was reduced in the brain and Nkx-2.5 expression was dramatically reduced in the heart of SRSF1 morphants. In addition, a dramatic reduction in functional chordin RNA in SRSF1 morphants was observed suggesting that chordin is one of the targets of SRSF1. Thus, we concluded that SRSF1 is an essential factor for pattern formation including heart, cartilage and germ lay PMID: 27396620
- results strongly support SRSF1 as a prognostic biomarker in SCLC and provide a rationale for personalized therapy in SCLC PMID: 27093186
- NSrp70 acts as a new molecular counterpart for alternative splicing of target RNA, counteracting SRSF1 and SRSF2 splicing activity. PMID: 26797131
- we have identified for the first time a potential mechanism that involves the essential splicing factor ASF/SF2 through which morphine regulates splicing specificity of the MOR encoding gene, OPRM1. PMID: 26553431
- Global splicing analysis with RNA-seq revealed that exons carrying the hnRNP H-binding GGGGG motif are predisposed to be skipped compared to those carrying the SRSF1-binding GGAGG motif in both human and mouse brains. PMID: 26282582
- We present experimental evidence that splicing factor SRSF1, SRSF2, U2AF35, U2AF65 and KHSRP expression levels in gastrointestinal tract (colon, gastric and pancreatic) tumors differ compare to healthy tissues and in cell lines PMID: 26406946
- These findings have revealed a molecular interplay between cellular SRSF1 and viral T-antigen in controlling JC virus gene expression. PMID: 26596376
- These include alternative splicing of SRSF1 itself. PMID: 26273603
- SF2 contributes to the elevated levels of ERK activation in hepatocellular carcinoma cells through modulating key component(s) downstream of growth factor receptors and upstream of ERK. PMID: 26018840
- These results suggest that MALAT1 may function as a promoter of gastric cancer cell proliferation partly by regulating SF2/ASF. PMID: 24857172
- SRSF1 regulates CD3zeta expression in human T cells and may contribute to the T cell defect in systemic lupus erythematosus. PMID: 26134847
- serine/arginine-rich splicing factor 1 (SRSF1) could specifically interact with RIG-I to facilitate RIG-I mediated production of type-I IFN that is triggered by cytosolic DNA. PMID: 25658361
- Using RNA-sequencing and 3-D cell cultures that mimic breast cancer, the authors identified SRSF1-regulated alternative splicing targets that are candidates for mediating SRSF1's oncogenic functions in mammary cells. PMID: 26431027
- The accumulated knowledge about SRSF1 provides critical insight into the integral role it plays in maintaining cellular homeostasis and suggests new targets for anticancer therapy. PMID: 24807918
- Data suggest that proline phosphorylation by CLK1/CDC-like kinase 1 (but not by SRPK1/serine/arginine-rich splicing factor kinase 1) regulates conformation and alternative splicing function of SFRS1 (serine/arginine-rich splicing factor 1). PMID: 25529026
- SRSF1 is a critical regulator that controls both RNA splicing and stability in the nucleus and functions in a Malat1-dependent manner. PMID: 24468535
- upon T cell activation, SRSF1 becomes limiting, and its function in CD6 exon 5 splicing is countered by an increase in CD6 transcription, dependent on chromatin acetylation. PMID: 24890719
- Data indicate that armadillo repeat protein ARVCF interacts with the splicing factors the splicing factor SRSF1 (SF2/ASF), the RNA helicase p68 (DDX5), and the heterogeneous nuclear ribonucleoprotein hnRNP H2. PMID: 24644279
- In the broader context of cancer pathology, the results suggest that SRSF1 might play a central role not only in the tumor cells, but also in the surrounding stroma. PMID: 23966470
- results demonstrate a novel mechanism of regulation of the splicing factor SRSF1 in human T cells and a potential molecular mechanism that controls its expression in SLE. PMID: 24368769
- Identification of new splicing events regulated by the oncogenic splicing factor SRSF1 in lung cancer using novel analytical tool. PMID: 24371231
- Specific induction of AS2 due to Alternative splicing is associated with epithelial ovarian cancer. PMID: 23748175
- Overexpression of SRSF1 and SRSF9 promote beta-catenin accumulation via the recruitment of beta-catenin mRNA and by enhancing its translation in an mTOR-dependent manner. PMID: 23592547
- The data establish a new view of SRSF1 protein regulation in which SRPK1 and CLK1 partition activities based on Ser-Pro versus Arg-Ser placement rather than on N- and C-terminal preferences along the RS domain. PMID: 23707382
- our results indicate that ASF/SF2 binds to a purine-rich region distant from both the previously published initiation site of HDV mRNA transcription and binding site of RNAP II, and suggest that this protein is not involved in HDV replication PMID: 23349975
- a new mechanism of posttranscriptional regulation of DIO1 and show deregulation of DIO1 expression in pituitary adenoma, possibly resulting from disturbed expression of SF2/ASF. PMID: 23462647
- High expression of ASF is associated with chronic myeloid leukemia. PMID: 23228155
- Results indicate that SF2/ASF regulates IL-2 production and that decreased SF2/ASF expression in systemic lupus erythematosus (SLE) T cells contributes to deficient IL-2 production. PMID: 23319613
- Results indicate that SRSF1 serves as an anti-apoptotic factor and potentially contributes to leukemogenesis in pediatric ALL patients by cooperating with PRMT1. PMID: 22839530
- Data indicate that depletion of SRSF1 in human cells compromises the association of splicing factors to nuclear speckles and influences the levels and activity of other SR proteins. PMID: 22855529
- SRSF1 is a direct target of the transcription factor oncoprotein MYC. PMID: 22545246
- Overexpression of SRSF1 rescues apoptosis of MCF-7 cells induced by Pnn depletion. PMID: 22454513
- In three-dimensional (3D) culture, SRSF1-overexpressing MCF-10A cells formed larger acini than control cells, reflecting increased proliferation and delayed apoptosis during acinar morphogenesis. PMID: 22245967
- ASF/SF2 translocates to the cytosol and regulates the alternative splicing of endoglin during senescence of endothelial cells. PMID: 21668763
- a novel intronic splicing enhancer that regulates caspase 9 RNA splicing and specifically interacts with SRSF1 was identified. PMID: 21622622
- negative regulation of the JC virus promoter by SF2/ASF may control reactivation of JCV replication in brain PMID: 21297941
- phosphorylation of the RS domain in SRSF1 induces a key molecular switch from intra- to intermolecular interactions, suggesting a plausible mechanism for the documented requirement for the phosphorylation/dephosphorylation cycle during pre-mRNA splicing. PMID: 21536904
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Nucleus speckle.
-
蛋白家族:Splicing factor SR family
-
數據庫鏈接:
Most popular with customers
-
Human Transforming Growth factor β1,TGF-β1 ELISA kit
Detect Range: 23.5 pg/ml-1500 pg/ml
Sensitivity: 5.8 pg/ml
-

-
-

Mouse Tumor necrosis factor α,TNF-α ELISA Kit
Detect Range: 7.8 pg/ml-500 pg/ml
Sensitivity: 1.95 pg/ml
-
-
-
-