Recombinant Mouse Stimulator of interferon genes protein (Sting1), partial
-
中文名稱:小鼠Sting1重組蛋白
-
貨號:CSB-EP023754MO1-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Sting1; Eris; Mita; Mpys; Tmem173; Stimulator of interferon genes protein; mSTING; Endoplasmic reticulum interferon stimulator; ERIS; Mediator of IRF3 activation; MMITA; Transmembrane protein 173
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Facilitator of innate immune signaling that acts as a sensor of cytosolic DNA from bacteria and viruses and promotes the production of type I interferon (IFN-alpha and IFN-beta). Innate immune response is triggered in response to non-CpG double-stranded DNA from viruses and bacteria delivered to the cytoplasm. Acts by binding cyclic dinucleotides: recognizes and binds cyclic di-GMP (c-di-GMP), a second messenger produced by bacteria, and cyclic GMP-AMP (cGAMP), a messenger produced by CGAS in response to DNA virus in the cytosol. Upon binding of c-di-GMP or cGAMP, STING1 oligomerizes, translocates from the endoplasmic reticulum and is phosphorylated by TBK1 on the pLxIS motif, leading to recruitment and subsequent activation of the transcription factor IRF3 to induce expression of type I interferon and exert a potent anti-viral state. In addition to promote the production of type I interferons, plays a direct role in autophagy. Following cGAMP-binding, STING1 buds from the endoplasmic reticulum into COPII vesicles, which then form the endoplasmic reticulum-Golgi intermediate compartment (ERGIC). The ERGIC serves as the membrane source for WIPI2 recruitment and LC3 lipidation, leading to formation of autophagosomes that target cytosolic DNA or DNA viruses for degradation by the lysosome. The autophagy- and interferon-inducing activities can be uncoupled and autophagy induction is independent of TBK1 phosphorylation. Autophagy is also triggered upon infection by bacteria: following c-di-GMP-binding, which is produced by live Gram-positive bacteria, promotes reticulophagy. Exhibits 2',3' phosphodiester linkage-specific ligand recognition: can bind both 2'-3' linked cGAMP (2'-3'-cGAMP) and 3'-3' linked cGAMP but is preferentially activated by 2'-3' linked cGAMP. The preference for 2'-3'-cGAMP, compared to other linkage isomers is probably due to the ligand itself, whichs adopts an organized free-ligand conformation that resembles the STING1-bound conformation and pays low energy costs in changing into the active conformation. May be involved in translocon function, the translocon possibly being able to influence the induction of type I interferons. May be involved in transduction of apoptotic signals via its association with the major histocompatibility complex class II (MHC-II).
-
基因功能參考文獻:
- Ubxn3b(-/-), like Sting(-/-) mice, are highly susceptible to lethal herpes simplex virus 1 (HSV-1) and vesicular stomatitis virus (VSV) infection, which is correlated with deficient immune responses when compared to Ubxn3b(+/+) littermates. PMID: 29899553
- this study shows that STING-/- mice presented defective protective mechanisms of intestinal mucosa, including decreased number of goblet cells, diminished mucus production, and lower levels of secretory IgA PMID: 29346345
- In response to the presence of cytosolic DNA, STING translocates from the endoplasmic reticulum (ER) to the Golgi, and activates TANK-binding kinase 1 (TBK1), a cytosolic kinase that is essential for the activation of STING-dependent downstream signaling. TBK1 binds to STING at the Golgi, not at the ER. PMID: 29870684
- Usp13 deconjugates polyubiquitin chains from STING and prevents the recruitment of Tbk1 to the signalling complex, thereby negatively regulating cellular antiviral responses. PMID: 28534493
- STING-mediated innate immune responses and dendritic cell maturation do not require TICAM-1 in myeloid lineage immune cells. PMID: 29627569
- PUMA promotes the cytosolic release of mitochondrial DNA and activation of the DNA sensors DAI/Zbp1 and STING, leading to enhanced RIP3 and MLKL phosphorylation in a positive feedback loop. PMID: 29581256
- identified nitro-fatty acids as endogenously formed inhibitors of STING signaling and propose for these lipids to be considered in the treatment of STING-dependent inflammatory diseases. PMID: 30061387
- Data show that mice defective in cyclic GMP-AMP synthase (cGAS) or STING protein (STING) are highly susceptible to acute herpes simplex encephalitis (HSE) . PMID: 27830700
- The STING/type I interferon pathway enhances suppressive inflammation in tumors by recruiting myeloid cells in part via the CCR2 pathway. Germ-line knockouts of CCR2 or treatment with an anti-CCR2 antibody results in blockade of radiation-induced MDSC infiltration. PMID: 29170400
- the induction of STING signaling is contingent on a fine-tuning of intracellular calcium levels. PMID: 29673589
- STING was dispensable for restricting S. pneumoniae during acute pneumonia in mice. PMID: 29263110
- data demonstrate that numerous RNA viruses evade cGAS/STING-dependent signaling and affirm the importance of this pathway in shaping the host range of ZIKV. PMID: 29915078
- Immune activation of STING requires palmitoylation at the Golgi. PMID: 27324217
- STING has dual functions in host defense, regulating protein synthesis to prevent RNA virus infection and regulating IFN expression to restrict DNA viruses PMID: 29440426
- The findings provide biochemical and imaging evidence for STING degradation by the lysosome and pinpoint trafficking-mediated STING degradation as a previously unanticipated therapeutic target for enhancing STING signaling in cancer therapy. PMID: 29241549
- The cGAS-STING cascade contributes to antibacterial defense against L. pneumophila in mice and men, and provides important insight into how the common HAQ TMEM173/STING variant affects antimicrobial immune responses and susceptibility to infection. PMID: 29298342
- DsbA-L prevents obesity-induced inflammation and insulin resistance by suppressing the mtDNA release-activated cGAS-cGAMP-STING pathway. PMID: 29087318
- Nontypeable Haemophilus influenzae DNA as a Pathogen-Associated Molecular Pattern Molecules triggered I-IFN response, which was STING/TBK1/IRF3 dependent. PMID: 29421524
- Intratumoral administration of the STING agonist cyclic di-GMP (CDG) or Flt3 Ligand (Flt3L) augmented the therapeutic effect of systemic triple checkpoint modulation and promoted the cure of 75% of mice with bilateral TRAMP-C2; however, when all agents were administered locally, only CDG mobilized abscopal immunity PMID: 28674082
- provide genetic evidence that cell-autonomous control of lentivirus infection in myeloid cells by SAMHD1 limits virus-induced production of interferons and the induction of co-stimulatory markers PMID: 27477283
- mouse primary T cells and T leukaemia are hyperresponsive to STING agonist, and this strong STING signalling is associated with apoptosis induction. PMID: 28874664
- STING-regulated pathways underlie the pathogenesis of many diseases including infectious diseases and cancers. It has also become evident from these studies that STING is a promising therapeutic target for the treatment of cancer. PMID: 26980676
- STING activated an antiviral/type I interferon response with live but not killed S. aureus. PMID: 28704551
- Data show that stimulator of interferon genes (STING)-associated vasculopathy with onset in infancy (SAVI)-associated STING N153S mutation triggers IRF3-independent immune cell dysregulation and lung disease in mice. PMID: 28951494
- These results highlight the crucial role of MFN1 in maintaining the competency of the STING pathway. PMID: 28729291
- Findings identify stress-mediated endoplasmic reticulum-phagy as a cell-autonomous response mobilized by STING-dependent sensing of a specific vita-pathogen-associated molecular patterns and elucidate how innate receptors engage multilayered homeostatic mechanisms to promote immunity and survival after infection. PMID: 29056340
- to contain the spread of herpes simplex virus type 1 in vivo, STING-dependent signaling leads to the upregulation of tetherin, a viral restriction factor PMID: 26627457
- study identifies the AIM2 inflammasome and cGAS/IFI16-STING-type I IFN pathway as a novel mechanism for host innate immunity to the ALVAC vaccine vector. PMID: 28947539
- NEMO was critically involved in the cGAS-STING pathway. PMID: 28939760
- found that CCCP impairs the interaction between STING and TBK1 and concomitantly triggers mitochondria fission. PMID: 28859978
- Yersinia YopJ negatively regulates IRF3-mediated antibacterial response through disruption of STING-mediated cytosolic DNA signaling. PMID: 27742471
- study provides evidence of STING activation in T cells, in which STING agonists not only provoke type I IFN production and IFN-stimulated gene expression, mirroring the response of innate cells, but are also capable of activating cell stress and death pathways. PMID: 28615418
- demonstrate that in colon tumor model, therapeutic anti-CD47 preferentially relies on STING-mediated DNA sensing in dendritic cells PMID: 28801234
- STING is a central mediator of interferon regulated inflammasome activation in Chlamydia trachomatis infection. PMID: 28570638
- TRIF and STING interacted directly, through their carboxy-terminal domains, to promote STING dimerization, intermembrane translocation, and signaling. PMID: 27631700
- We conclude that the R71H-G230A-R293Q (HAQ) of TMEM173 is a null TMEM173 allele. PMID: 27927967
- results show that resistance to HSV-1 in the trigeminal ganglia during acute infection is conferred in part by STING and IFN-alpha/beta signaling in both bone marrow-derived and -resident cells, which coalesce to support a robust HSV-1-specific CD8(+) T cell response PMID: 27511736
- results suggest that LSm14A plays an important role in antiviral innate and adaptive immune responses by modulating MITA level in a cell type-specific manner PMID: 27183626
- STING and TRIF Contribute to Mouse Sepsis, Depending on Severity of the Disease Model PMID: 27755506
- Data suggest that activation of either RIG-I/MAVS or STING pathways during acute intestinal tissue injury in mice resulted in IFN-I signaling that maintained gut epithelial barrier integrity and reduced GVHD severity. PMID: 28424327
- this study shows that iRhom2 is essential for STING activity, as it regulates TRAPbeta-mediated translocation and EIF3S5-mediated deubiquitination of STING PMID: 27428826
- The mitochondrial damage-cGAS-STING-IRF3 pathway is critically involved in metabolic stress-induced endothelial inflammation. PMID: 28302626
- Data show that STING induces tumor cytotoxicity by NK cells through tumor and host immune cell network to contribute to innate surveillance and suppression of tumors in vivo. PMID: 27608599
- Results reveal a greater complexity in the role of STING signaling in cancer, underscoring how innate immune pathways in the TME modify tumorigenesis in distinct tumor settings. PMID: 26964621
- NLRX1 sequesters the DNA-sensing adaptor STING from interaction with TBK1, which is a requisite for IFN-1 induction in response to viral DNA. PMID: 27078069
- Sting protein role in the innate immune response and interferon-stimulated genes response pathway PMID: 26903602
- required for chitosan-mediated enhancement of antigen specific Th1 and immunoglobulin G2 responses following vaccination PMID: 26944200
- in vitro activation of the AIM2 inflammasome in murine macrophages and dendritic cells leads to reduced activation of the STING pathway, in part through promoting caspase-1-dependent cell death. PMID: 26927800
- A STING-dependent, cGAS-independent pathway important for full interferon production and antiviral control of enveloped RNA viruses. PMID: 26893169
- These data provide evidence that the N-terminal domain of STING affects DNA responses via control of trafficking PMID: 26685207
顯示更多
收起更多
-
亞細胞定位:Endoplasmic reticulum membrane; Multi-pass membrane protein. Cytoplasm, perinuclear region. Endoplasmic reticulum-Golgi intermediate compartment membrane; Multi-pass membrane protein. Cytoplasmic vesicle, autophagosome membrane; Multi-pass membrane protein. Mitochondrion outer membrane; Multi-pass membrane protein. Cell membrane; Multi-pass membrane protein.
-
蛋白家族:TMEM173 family
-
組織特異性:Present in spleen and thymus tissue. Also present in dendritic cells (at protein level).
-
數據庫鏈接:
Most popular with customers
-
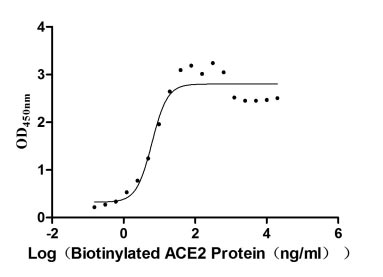
Recombinant Human Angiotensin-converting enzyme 2 (ACE2), partial,Biotinylated (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
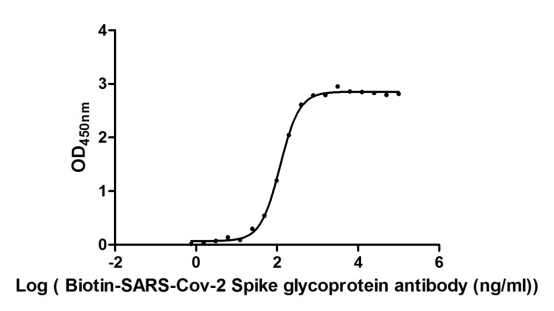
Recombinant Severe acute respiratory syndrome coronavirus 2 Spike glycoprotein (S), partial (Active)
Express system: Mammalian cell
Species: Severe acute respiratory syndrome coronavirus 2 (2019-nCoV) (SARS-CoV-2)
-
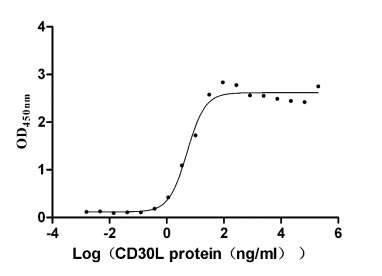
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human papillomavirus type 16 Protein E7 (E7) (Active)
Express system: E.coli
Species: Human papillomavirus type 16
-
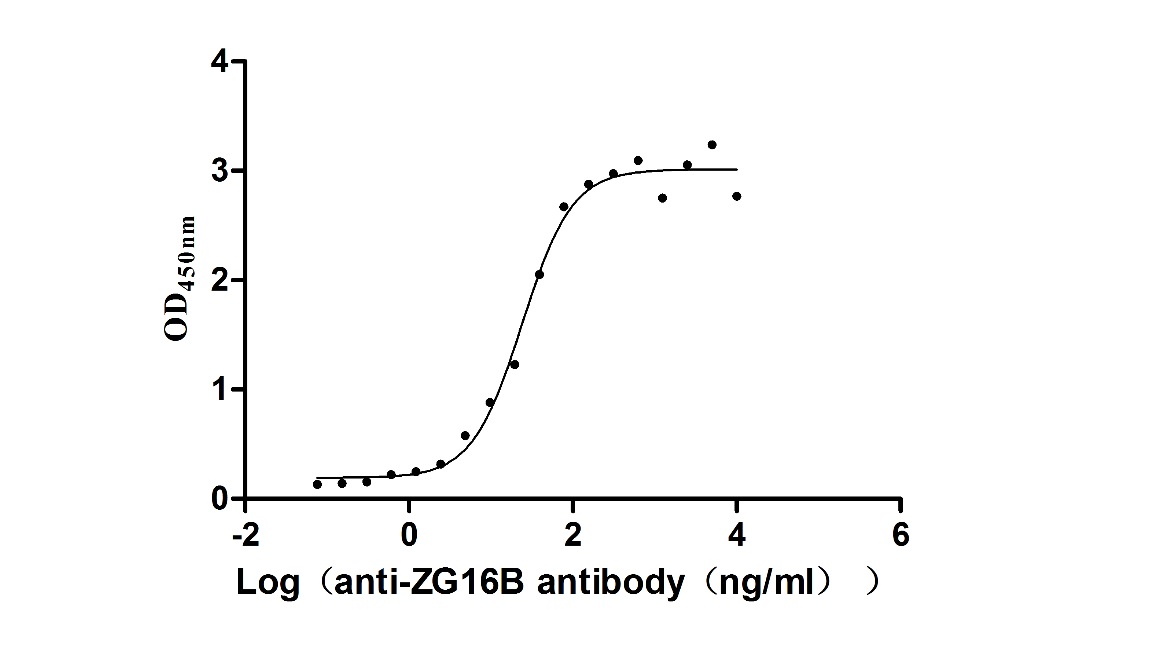
Recombinant Macaca fascicularis zymogen granule protein 16 homolog B (ZG16B) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
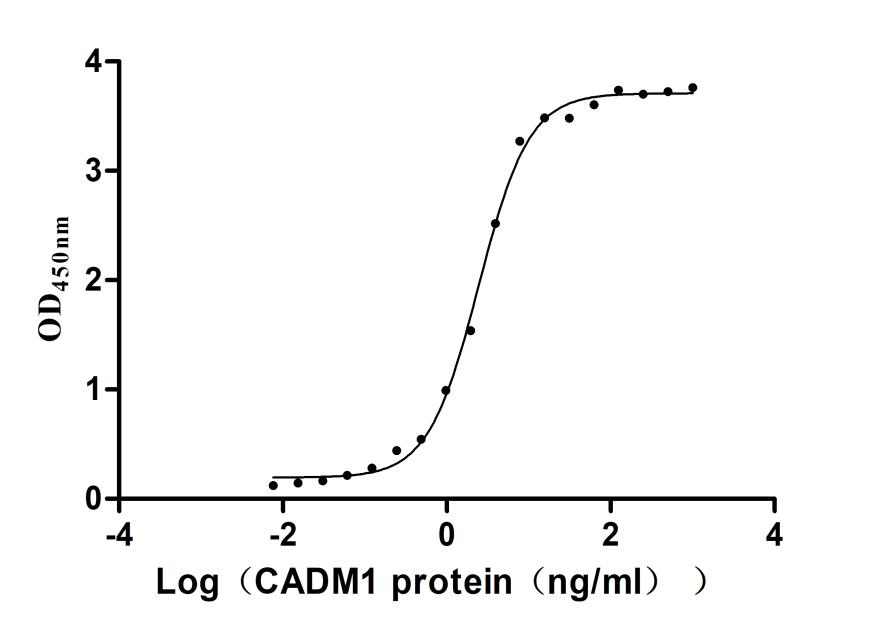
Recombinant Human Cytotoxic and regulatory T-cell molecule (CRTAM), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
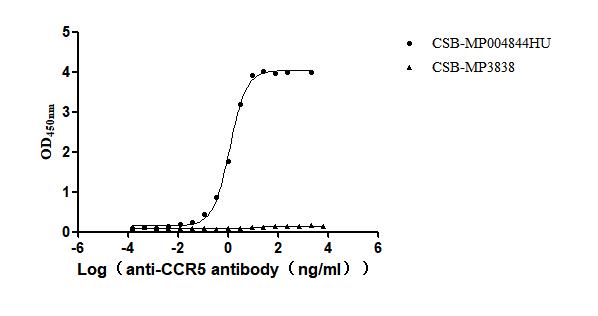
Recombinant Human C-C chemokine receptor type 5 (CCR5)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)










