Recombinant Mouse Serine/arginine-rich splicing factor 1 (Srsf1)
-
中文名稱:小鼠Srsf1重組蛋白
-
貨號:CSB-EP744112MO-B
-
說明書:
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Srsf1; Sfrs1; Serine/arginine-rich splicing factor 1; ASF/SF2; Pre-mRNA-splicing factor SRp30a; Splicing factor; arginine/serine-rich 1
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
表達區(qū)域:2-248
-
氨基酸序列SGGGVIRGP AGNNDCRIYV GNLPPDIRTK DIEDVFYKYG AIRDIDLKNR RGGPPFAFVE FEDPRDAEDA VYGRDGYDYD GYRLRVEFPR SGRGTGRGGG GGGGGGAPRG RYGPPSRRSE NRVVVSGLPP SGSWQDLKDH MREAGDVCYA DVYRDGTGVV EFVRKEDMTY AVRKLDNTKF RSHEGETAYI RVKVDGPRSP SYGRSRSRSR SRSRSRSRSN SRSRSYSPRR SRGSPRYSPR HSRSRSRT
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評價

靶點詳情
-
功能:Plays a role in preventing exon skipping, ensuring the accuracy of splicing and regulating alternative splicing. Interacts with other spliceosomal components, via the RS domains, to form a bridge between the 5'- and 3'-splice site binding components, U1 snRNP and U2AF. Can stimulate binding of U1 snRNP to a 5'-splice site-containing pre-mRNA. Binds to purine-rich RNA sequences, either the octamer, 5'-RGAAGAAC-3' (r=A or G) or the decamers, AGGACAGAGC/AGGACGAAGC. Binds preferentially to the 5'-CGAGGCG-3' motif in vitro. Three copies of the octamer constitute a powerful splicing enhancer in vitro, the ASF/SF2 splicing enhancer (ASE) which can specifically activate ASE-dependent splicing. Specifically regulates alternative splicing of cardiac isoforms of CAMK2D, LDB3/CYPHER and TNNT2/CTNT during heart remodeling at the juvenile to adult transition. The inappropriate accumulation of a neonatal and neuronal isoform of CAMKD2 in the adult heart results in aberrant calcium handling and defective excitation-contraction coupling in cardiomyocytes. May function as export adapter involved in mRNA nuclear export through the TAP/NXF1 pathway.
-
基因功能參考文獻:
- SRSF1 promotes vascular smooth muscle cell (VSMC) proliferation and injury-induced neointima formation. SRSF1 favors the induction of a truncated p53 isoform, Delta133p53, which has an equal proliferative effect and in turn transcriptionally activates Kruppel-like factor 5 (KLF5) via the Delta133p53-EGR1 complex, resulting in an accelerated cell-cycle progression and increased VSMC proliferation. PMID: 28799539
- In addition, overexpression of SRSF1 in XRCC4-deficient cells restored the normal level of apoptosis, suggesting that SRSF1 functions downstream of XRCC4 in activating CAD. PMID: 29233683
- SRSF1 is a key regulator of DBF4B pre-mRNA splicing dysregulation in colon cancer. SRSF1 is required for cancer cell proliferation. PMID: 29262322
- Authors showed that Mir505-3p was capable of inhibiting tumor proliferation driven by SRSF1 in two neural tumor cell lines, Neuro-2a (N2a) and U251, exclusively in serum-reduced condition. Authors observed that the protein level of SRSF1 was gradually promoted by increasing concentration of serum. PMID: 29120871
- This study showed that the splicing factor kinase SRPK1 is a key regulator of spinal nociceptive processing in naive and nerve injured animals. We present evidence for a novel mechanism in which altered SRSF1 localization/function in neuropathic pain results in sensitization of spinal cord neurons. PMID: 27616424
- The expression levels of three splicing factors, ESRP1, PTB and SF2/ASF, are significantly altered during cardiac hypertrophy in mice. PMID: 24552714
- Deletion of RRM1 eliminated the splicing activity of SRSF1 and thus cellular transformation. PMID: 23843040
- Specific effects on regulated splicing by SR proteins SRSF1 and SRSF2 depends on a complex set of relationships with multiple other SR proteins in mammalian genomes. PMID: 23562324
- Treatment with IL-17 prolongs the half-life of chemokine CXCL1 mRNA via the adaptor TRAF5 and the splicing-regulatory factor SF2 (ASF). PMID: 21822258
- analysis of the miRNA-mediated interaction between leukemia/lymphoma-related factor (LRF) and alternative splicing factor/splicing factor 2 (ASF/SF2) affects cell senescence and apoptosis PMID: 20923760
- Modulation of Xist RNA processing may be part of the stochastic process that determines which X chromosome will be inactivated. PMID: 20657585
- Disruption of an SF2/ASF-dependent exonic splicing enhancer in SMN2 causes spinal muscular atrophy in the absence of SMN1 PMID: 11925564
- Both hnRNP A1 and alternative splicing factor/splicing factor 2 contents rose in adenomas and during injury-induced hyperplasia compared to control lungs PMID: 15390079
- These results highlight the requirement of Sfrs1-mediated alternative splicing for the survival of retinal neurons, with sensitivity defined by the window of time in which the neuron was generated. PMID: 18987029
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Nucleus speckle.
-
蛋白家族:Splicing factor SR family
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-

Recombinant Human Insulin growth factor-like family member 1 (IGFL1) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
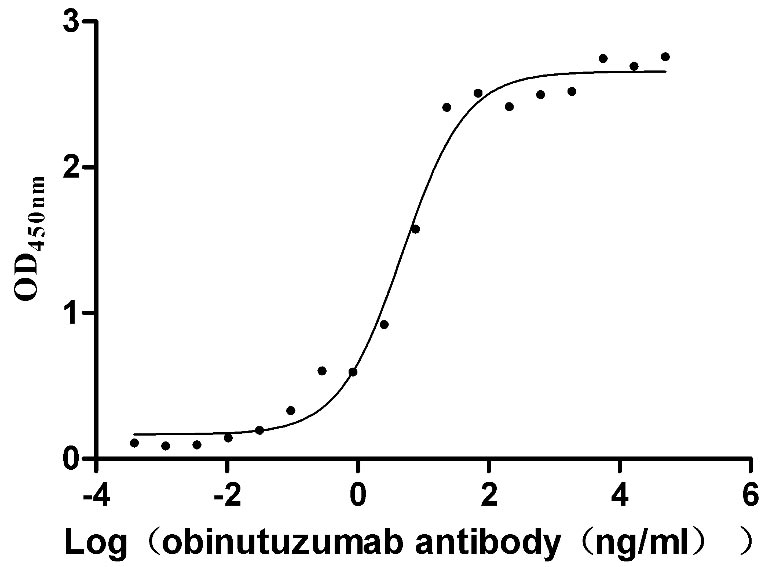
Recombinant Human B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
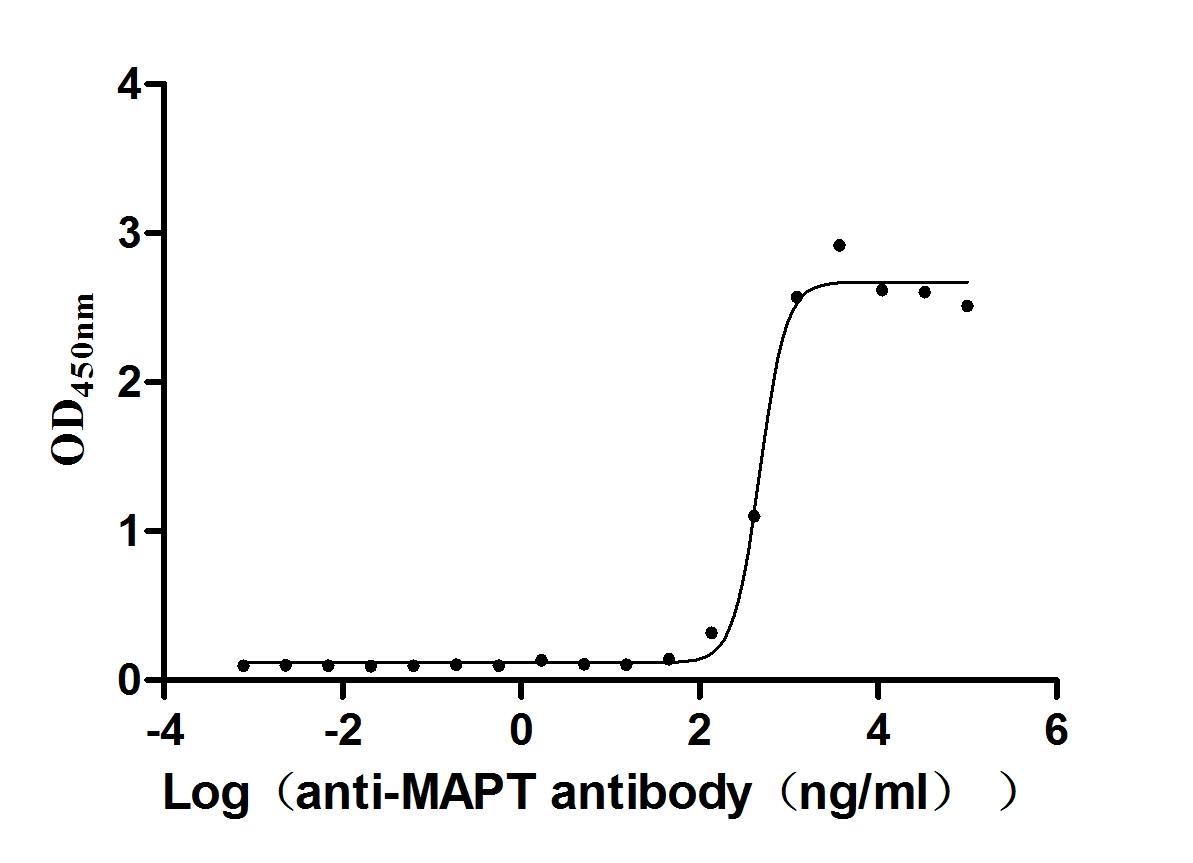
Recombinant Mouse Microtubule-associated protein tau (Mapt) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
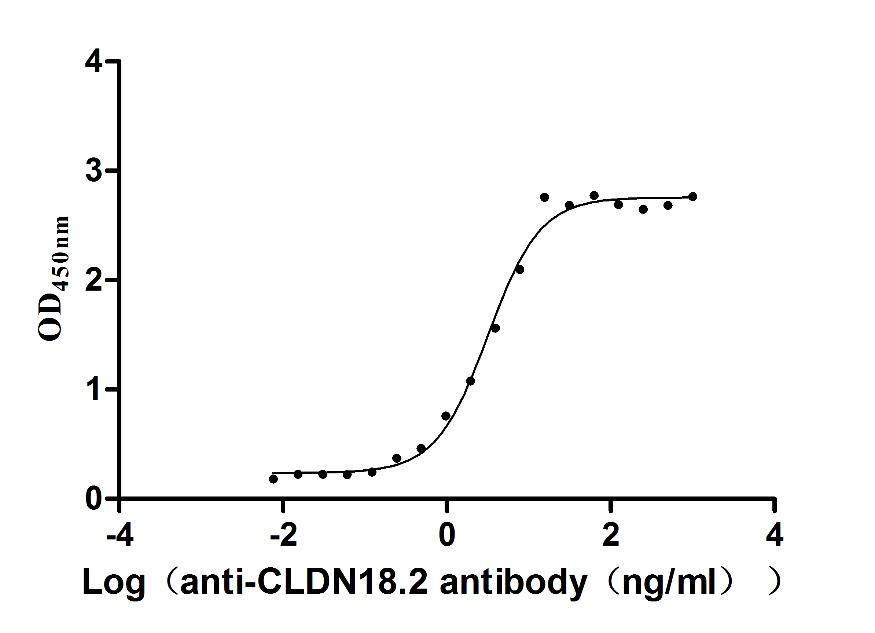
Recombinant Macaca fascicularis Claudin 18.2 (CLDN18.2)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Mouse Complement component C1q receptor (Cd93), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Human Glucagon-like peptide 1 receptor (GLP1R), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Claudin-3 (CLDN3)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human CUB domain-containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)










