RARRES3 Antibody, Biotin conjugated
-
中文名稱:RARRES3兔多克隆抗體, Biotin偶聯
-
貨號:CSB-PA892340LD01HU
-
規格:¥880
-
其他:
產品詳情
-
產品名稱:Rabbit anti-Homo sapiens (Human) RARRES3 Polyclonal antibody
-
Uniprot No.:
-
基因名:RARRES3
-
別名:HRASLS4 antibody; MGC8906 antibody; RAR responsive protein TIG3 antibody; RAR-responsive protein TIG3 antibody; RARRES3 antibody; Retinic acid inducible gene 1 antibody; Retinoic acid receptor responder (tazarotene induced) 3 antibody; Retinoic acid receptor responder protein 3 antibody; Retinoid inducible gene 1 protein antibody; Retinoid-inducible gene 1 protein antibody; RIG 1 antibody; rig1 antibody; Tazarotene induced gene 3 protein antibody; Tazarotene-induced gene 3 protein antibody; TIG 3 antibody; TIG3 antibody; TIG3_HUMAN antibody
-
宿主:Rabbit
-
反應種屬:Human
-
免疫原:Recombinant Human Retinoic acid receptor responder protein 3 protein (1-133AA)
-
免疫原種屬:Homo sapiens (Human)
-
標記方式:Biotin
-
克隆類型:Polyclonal
-
抗體亞型:IgG
-
純化方式:>95%, Protein G purified
-
濃度:It differs from different batches. Please contact us to confirm it.
-
保存緩沖液:Preservative: 0.03% Proclin 300
Constituents: 50% Glycerol, 0.01M PBS, PH 7.4 -
產品提供形式:Liquid
-
應用范圍:ELISA
-
Protocols:
-
儲存條件:Upon receipt, store at -20°C or -80°C. Avoid repeated freeze.
-
貨期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time maybe differs from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
-
用途:For Research Use Only. Not for use in diagnostic or therapeutic procedures.
產品評價

相關產品
靶點詳情
-
功能:Exhibits both phospholipase A1/2 and acyltransferase activities. Shows phospholipase A1 (PLA1) and A2 (PLA2), catalyzing the calcium-independent release of fatty acids from the sn-1 or sn-2 position of glycerophospholipids. For most substrates, PLA1 activity is much higher than PLA2 activity. Shows O-acyltransferase activity, catalyzing the transfer of a fatty acyl group from glycerophospholipid to the hydroxyl group of lysophospholipid. Shows N-acyltransferase activity, catalyzing the calcium-independent transfer of a fatty acyl group at the sn-1 position of phosphatidylcholine (PC) and other glycerophospholipids to the primary amine of phosphatidylethanolamine (PE), forming N-acylphosphatidylethanolamine (NAPE), which serves as precursor for N-acylethanolamines (NAEs). Promotes keratinocyte differentiation via activation of TGM1.
-
基因功能參考文獻:
- RIG-I and MDA5 receptor sensing of host non-coding RNAs facilitates the cell-intrinsic immune response to Kaposi's sarcoma-associated herpesvirus infection. PMID: 30451863
- we observed frameshift mutations of RARRES3 in 11 cases of colorectal cancer PMID: 29496306
- Data demonstrate that RIG-1 is activated after mitochondrial SUMOylation by MAPL which is an essential process for the mitochondrial antiviral signaling response. PMID: 28273895
- Low RIG1 expression is associated with Arenaviral infections. PMID: 29669840
- Conclusively, these data demonstrate the MCPIP1 contributes to attenuate influenza A virus-induced host antiviral response by suppressing RIG-I expression. PMID: 28892164
- Oncostatin M induces RIG-I and MDA5 expression and enhances the double-stranded RNA response in fibroblasts. PMID: 28560754
- Identified the tumor suppressor RARRES3 as a critical target of G9a. Epigenetic silencing of RARRES3 contributed to the tumor-promoting function of G9a PMID: 28532996
- Results provide evidence that RIG-I as an essential mediator for influenza A virus-induced COX-2 expression. PMID: 27265729
- perifascicular pattern of RIG-I expression supports the diagnosis of dermatomyositis PMID: 28738907
- The current study establishes that hypoxia can profoundly influence the inducible RIG-I protein expression in malignant cells of both human and murine origin, whereas this phenomenon was not identified in nonmalignant cell lines or primary cells. PMID: 28468914
- our study demonstrates that the novel pathway lncRNA Ftx/miR-545/RIG-I promotes hepatocellular carcinoma development PMID: 26992218
- this review describes antiviral activities of RIG-I against influenza viruses (standing on three legs) PMID: 27318973
- Study uncovered a novel aspect of Rig-I in monitoring gut microbiota through regulating IgA and IL6-STAT3-dependent Reg3gamma pathway. Besides, Rig-I loss could also promote colorectal cancer progression both in the presence and absence of intestinal bacteria. PMID: 28057020
- a novel mechanism of pattern recognition receptor modulation by HCV and suggests a biological function of the HCV alternate reading frame in the modulation of host innate immunity PMID: 27404108
- this study indicated that increased expression of TIG3 in primary glioblastoma is a novel biomarker for predicting poor outcome of patients. We then hypothesize that TIG3 may function in a different pattern in glioblastoma PMID: 28639915
- Foot-and-mouth disease virus Viroporin 2B antagonizes RIG-I-mediated antiviral effects by inhibition of its protein expression. PMID: 27707918
- Overexpression of TIG3 suppresses tumor growth in hepatocellular carcinoma PMID: 26951515
- miR-34a is an antioncogene in multiple tumors, in this study, RIG-I and miR-34a suppressed cell growth, proliferation, migration, and invasion in cervical cancer cells in vitro. miR-34a was validated as a new regulator of RIG-I by binding to its 3' untranslated region and upregulating its expression level. PMID: 26910120
- TRIM25 plays an additional role in RIG-I/MDA5 signaling other than RIG-I ubiquitination via activation of NF-kappaB. PMID: 26299329
- These observations highlight the importance of RIG-I signaling in anti-HIV innate immunity in macrophages, which may be beneficial for the treatment of HIV and aid in the understanding of the neuropathogenesis of HAND. PMID: 26535695
- Gal1 and gal3 regulate the inflammatory response in airway epithelial cells exposed to microbial neuraminidase by modulating the expression of SOCS1 and RIG1. PMID: 26355912
- findings provide a new insight into the molecular link between p53, protein acylation and Wnt/beta-catenin signaling whereby RARRES3 plays a pivotal role in modulating the acylation status of signaling proteins. PMID: 25361079
- During Crimean-Congo hemorrhagic fever virus infection, RIG-I mediated a type I interferon response via MAVS. PMID: 26223644
- the flexible main loop of H-REV107, but not that of TIG3, is critical for its NTD to modulate its CTD in inducing cell death. PMID: 25871522
- Viral pseudo-enzymes activate RIG-I via deamidation to evade cytokine production. PMID: 25752576
- critical for the initiation of IRF-3 phosphorylation, dimerization and downstream gene expression during antiviral innate immune response PMID: 24800889
- RARRES3 downregulation engages metastasis-initiating capabilities by facilitating adhesion of the tumor cells to the lung parenchyma. PMID: 24867881
- MiR-545 exerts its effects in pancreatic ductal adenocarcinoma by directly targeting RIG-1 PMID: 25315416
- These findings argue that TIG3 is involved in the control of keratinocyte differentiation and that loss of TIG3 in transformed cells contributes to the malignant phenotype. PMID: 24599174
- We show that the C-terminal hydrophobic domain targets intact TIG3 to the plasma membrane, but when isolated as an independent element localizes at the mitochondria and a segment of the N-terminal hydrophilic region targets the centrosome. PMID: 24401997
- Low RIG1 expression is associated with testis cancer. PMID: 22960220
- RIG1 gene is a downstream target of p53 in cancer cell lines PMID: 22616991
- Data suggest that pericentrosomal localization of TIG3 is a key event that results in microtubule and microfilament redistribution and pericentrosomal organelle clustering and that leads to cancer cell apoptosis. PMID: 21858038
- We conclude that TG1-catalysed cross-linking, regulated by TIG3, might play an important role in the formation of neuronal tau inclusions in certain tauopathies PMID: 22009441
- induction of TIG3 is associated with the suppression of anchorage-independent growth in certain aerodigestive tract cancer cells PMID: 12879006
- TIG3 facilitates the terminal stages in keratinocyte differentiation via activation of type I transglutaminase PMID: 12928434
- altered RARRES3 expression may play a role in the carcinogenesis of the liver and biliary tract. PMID: 15742394
- identification and characterization of the functional RA response element that is responsible for the RA-mediated induction of RIG1 gene PMID: 15850806
- TIG3 is negatively regulated by an activated MEK-ERK signaling pathway in ovarian carcinoma. PMID: 15856468
- The proapoptotic and anti-RAS activities of RIG1 are primarily associated with the Golgi localization of the protein. PMID: 17196792
- the TIG3 N-terminal region is required for TIG3-dependent keratinocyte differentiation, and its removal converts TIG3 into a proapoptotic protein PMID: 17762858
- The NC domain, especially the NC motif, plays the major role in RIG1-mediated pro-apoptotic activity. The RIG1(111-123) dodecapeptide exhibited strong pro-apoptotic activity and has potential as an anticancer drug. PMID: 19245694
- The tumor suppressors TIG3, HRASLS2 and H-rev107 are involved in the phospholipid metabolism with different physiological roles. PMID: 19615464
顯示更多
收起更多
-
亞細胞定位:Membrane; Single-pass membrane protein.
-
蛋白家族:H-rev107 family
-
組織特異性:Widely expressed.
-
數據庫鏈接:
Most popular with customers
-

-
Phospho-YAP1 (S127) Recombinant Monoclonal Antibody
Applications: ELISA, WB, IHC
Species Reactivity: Human
-
-
-
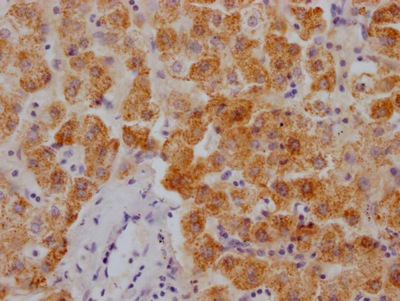
-
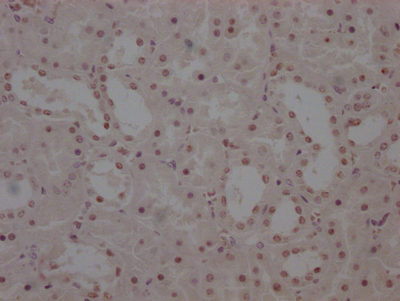
-
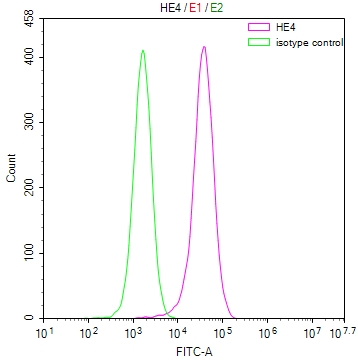
-